How can one ignore a book with a provocative title like “What Darwin got wrong”? Jerry Fodor and Massimo Piattelli-Palmarini argue that natural selection cannot be the driving force behind evolution. So here we have a book challenging the core of evolutionary biology, written bytwo cognitive scientists. It would seem like a futile exercise and unfortunately for the authors that is precisely what it is.
The authors make comparisons between the theory of natural selection and Skinner's theory of learning by operant conditioning. Fodor and Piattelli-Palmarini are after all cognitive scientists and cognitive psychology really took over from behaviorism and Skinner's theory of operant conditioning in the late 1950s and the 1960s. Operant conditioning theory describes modification of behavior as a result of learning associations between responses and reward or punishment. While it cannot be denied that operant conditioning provides a useful model for some forms of learning it is believed that behaviorists like Skinner took their theory too far and focused too much on stimuli and responses and ignored what happens in between, the mental states and cognitive processes that cognitive psychology holds so dearly. This is behaviorisms infamous back box. Fodor and Piattelli-Palmarini seem to be making the comparison between operant conditioning and natural selection, because they want to imply that natural selection essentially has the same kind of problems as operant conditioning. The whole comparison is really quite irrelevant; the theory of natural selection must be judged on its own merits.
The next step in Fodor and Piattelli-Palmarini's approach is to go through a number of developments in evolutionary biology, which they believe challenge Darwinism or neo-Darwinism (the synthesis of natural selection theory with Mendellian genetics). Among the developments discussed by the authors are epigenetics and evo-devo. I will focus on these developments here, but I will not deal with Fodor and Piattelli-Palmarini's arguments directly. In this section of their book they appear to be challenging a simple uni-dimensional evolutionary model, but this is not the essence of Darwinism. To challenge Darwinism one needs to show that Darwin was wrong about (1) descent with modification, or (2) natural selection as the main driving force behind evolution. I will argue that evo-devo and epigenetics pose no problem for Darwinism and natural selection. Fodor and Piattelli-Palmarini have a more direct assault against Darwinism in the remainder of their book and I will leave that for a later post.
Evo-devo, or evolutionary development, focuses on how genetic and epigenetic factors interact during the development of an organism. Epigenetics here simply refers to non-genetic (epi = besides, above) factors, or rather environmental factors that influence how a genome is expressed and how an organism develops into its phenotype.
In this picture, based on a photographic survey by Michael Richardson (see
PZ Myers blog), we see that there is remarkable similarity across vertebrate species in the phylotypic stage of development (top row). In this stage the basis for each subsequent vertebrate body plan is set. PZ Myers explains on his blog:
At this time in development, vertebrate embryos all express a suite of characters that are common to the entire vertebrate lineage: they have a notochord and a dorsal nerve cord, they have pharyngeal arches and a tail, and they have a repeating series of blocks of muscle called somites. Most of the features that distinguish different vertebrate groups, such as limbs or fins, hair or feathers or scales, and wings or forearms, haven't yet developed. In addition, we now know that this is the period during which a set of crucial pattern forming genes (the emx/otx/hox genes) is first expressed, and lay down the molecular blueprint of the body plan.
What evo-devo shows is that there are complex interactions between genes and that the effect of a certain mutation on the resulting phenotype is likewise complex. The simple unidimensional picture of a gene for x and a gene for y is to some extent misleading. Fodor and Piattelli-Palmarini are correct on this point. Does this in any way challenge Darwinism? Well, lets discuss two different ways in which development occurs. First, there's what is called canalized development, where a certain developmental trajectory becomes strongly regulated, so that is becomes less vulnerable to change. The activity of some genes can compensate for variability in other genes. Within such canalized developmental trajectories it can therefore occur that certain mutations of genes do not ultimately affect the phenotype. Why would some developmental trajectories become canalized? Ken Richardson explains in his book
The evolution of intelligent systems:
Many environmental conditions are sufficiently constant from generation to generation to allow the development of fairly uniform structures in organisms, with little variation between individuals. This is the classical Darwinian natural selection scenario, and the beaks of Darwin's finches are good examples, along with numerous other aspects of all living things......... canalization has evolved for developmental adaptation to aspects of environment that, like seeds for finches, reliably occur across generations.
Canalization is completely understandable from a Darwinian natural selection perspective. At the opposite extreme from canalization we have developmental plasticity. This is where epigenetics comes in. It turns out that there are environmental influences on certain developmental trajectories. Because of this the same genotype does not necessarily result in the same phenotype. There are many interesting examples of how epigenetics influences development and one of these was presented to me as I was working on this blog post. While watching a recorded episode of the BBC program QI I saw a picture of the first cloned cat:

The cloned cat CC (for copycat) is in the top right picture and in the bottom picture together with its surrogate mother. The genetic donor is in the top left picture. As you can see the coloration is different (compare the top two pictures). So we have the same genotype, but a different phenotype. Unfortunately QI got their facts wrong as they only showed the bottom picture (clone plus surrogate mother) and claimed they were genetically identical. Getting something like CC from a tabby seems a bit too much to ask for epigenetic effects.
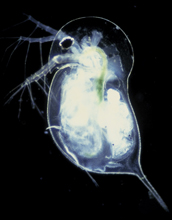
Epigenetic effects are seen in natural circumstances across the board. Lets start small and work our way up to humans. Richardson gives the example of the water flea Daphnia. If developing juveniles detect the presence of predatory midge larvae they develop protective neck spines or helmets. Interestingly, Daphnia with large helmets also produce offspring with helmets even in the absence of the midge larvae. The epigenetic effects are here transgenerational!
In the top picture we see a Daphnia with a clonal brood of offspring (i.e. genetically identical); in the bottom picture the Daphnia has developed a protective helmet (pictures from
the NSF).
Epigenetic factors also influence the timing of life history events in amphibians (metamorphosis) and even mammals (birth). For example,
Crespi and Denver (2005) found that tadpoles of the western spadefoot toad accelerate metamorphosis in response to water volume reduction or food restriction. The accelerated metamorphosis allows the toad to leave the aquatic environment so that it can find a more desirable environment, although this comes with a cost to its growth.
Crespi and Denver note that stressors such as maternal malnutrition, hypoxia or infection have effects in mammals that correspond to the accelerated development and early metamorphosis in amphibians. That is, fetal development is accelerated and the probability of preterm birth is dramatically increased. Humans are no exceptions. The 1944-1945 famine in the Netherlands caused many preterm babies and babies with lower birth weight. Maternal malnutrition also gave the children an increased chance of developing obesity and anti-social behavior. Even though these children did not experience malnutrition during their adult lives their children also turned out to have a lower birth weight. In other words, the epigenetic effects were to some extent transgenerational, just as the Daphnia protective helmets and neck spines.
Why do organisms respond to these external influences in these ways? Clearly, somehow the development is affected by chemicals, hormones and other signals that provide information about the environment. What determines how the organism responds to these signals? The answer here is natural selection. Far from being challenged by these findings, they make perfect sense from a Darwinian perspective. Chemicals and hormones from the external environment will interact with gene expressions in complex ways and the effects will naturally depend on which genes are being expressed. If there is a genetic mutation the external signals may have different effects. If the modified effect of the external signals somehow has a survival value then it will be favored by natural selection. It is of course no coincidence that Daphnia respond to chemical hormones of the predatory midge larvae by developing defense structures. This development was primed by natural selection. It is also no coincidence that tadpoles respond to low water levels or lack of nutrition by doing precisely what it should do to escape the pond, that is accelerate metamorphosis. Again this response must have been primed by natural selection. Finally, concerning the effects of maternal malnutrition during the Dutch famine of 1944-1945 it cannot be a coincidence that the effects on the developing fetus prepare the fetus for life outside the womb. These children favored fatty foods, precisely as you would expect if the child has been developmentally prepared for an environment in which food is scarce. Unfortunately this leads to obesity if it turns out that there actually is sufficient food in the environment. According to
Dick Swaab the anti-social behavior might also be part of the preparation for an environment in which one needs to struggle to get sufficient food. How is it possible that the fetus prepares for the environmental conditions? Dick Swaab explains (my translation):
To conclude, the fetus appears to “think” only about short-term survival and adjusts to the difficult circumstances that it expects immediately after birth. Of course, the fetus doesn't “think” about these things. Millions of years unborn children have been exposed to these kinds of threats. One time a child had a mutation that allowed it to adjust better to the problems that were waiting for it, and this favorable mutation spread through the population.
I would suspect that there were multiple mutations involved and that some of them might even go back tens if not hundreds of millions of years, given that there are similarities in the way amphibians and mammals respond to environmental factors.
Epigenetics, and the field of evo-devo in general, do not appear to challenge the theory of natural selection one bit, regardless of what Fodor and Piattelli-Palmarini claim. Apart from the fact that epigenetic effects are just as much subject to natural selection as genetic effects, it should also be noted that the transgenerational effects of epigenetics generally appear to be limited. Authors report that the effects can last for a few generations, but how does that compare to genetic mutations, some of which have been with us for over hundreds of millions of years? Perhaps the limited transgenerational effects are to be expected as these epigenetic effects seem very suitable for modifications contingent on the current situation in the environment. They are part of the developmental plasticity that is needed for changing environmental conditions.
Although there have been many interesting findings related to epigenetics it appears that not all evolutionary biologists are impressed. In
The greatest show on earth Richard Dawkins calls epigenetics “a modish buzz-word now enjoying its fifteen minutes of fame in the biological community." He also complains that its enthousiasts cannot even agree with themselves, let alone each other, what epigenetics actually means. Ouch! Here I've been using the term epigenetics as non-genetic effects during development that change the phenotype. It remains to be seen how the field of evo-devo will change evolutionary biology and what will become of epigenetics, but one trend certainly seems to be a greater appreciation for how organisms develop and how genes and environment interact in complicated ways to form a phenotype. My gut feeling is that an increased focus on evolutionary development is one good thing that has come out of this research.